Recombinant Mouse Interleukin-18 (Il18)
In Stock-
中文名稱:小鼠Il18重組蛋白
-
品名簡稱:Recombinant Mouse Il18 protein
-
貨號:CSB-YP011608MO
-
說明書:
-
規格:¥1836
-
圖片:
-
 (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
-
-
其他:
產品詳情
-
純度:Greater than 90% as determined by SDS-PAGE.
-
基因名:
-
Uniprot No.:
-
別名:Il18; Igif; Interleukin-18; IL-18; Interferon gamma-inducing factor; IFN-gamma-inducing factor; Interleukin-1 gamma; IL-1 gamma
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Full Length
-
來源:Yeast
-
分子量:24.1kDa
-
表達區域:1-192aa
-
氨基酸序列MAAMSEDSCVNFKEMMFIDNTLYFIPEENGDLESDNFGRLHCTTAVIRNINDQVLFVDKRQPVFEDMTDIDQSASEPQTRLIIYMYKDSEVRGLAVTLSVKDSKMSTLSCKNKIISFEEMDPPENIDDIQSDLIFFQKRVPGHNKMEFESSLYEGHFLACQKEDDAFKLILKKKDENGDKSVMFTLTNLHQS
Note: The complete sequence may include tag sequence, target protein sequence, linker sequence and extra sequence that is translated with the protein sequence for the purpose(s) of secretion, stability, solubility, etc.
If the exact amino acid sequence of this recombinant protein is critical to your application, please explicitly request the full and complete sequence of this protein before ordering. -
蛋白標簽:C-terminal 6xHis-tagged
-
產品提供形式:Liquid or Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
緩沖液:Tris-based buffer,50% glycerol
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:3-7 business days
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet & COA:Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:A proinflammatory cytokine primarily involved in polarized T-helper 1 (Th1) cell and natural killer (NK) cell immune responses. Upon binding to IL18R1 and IL18RAP, forms a signaling ternary complex which activates NF-kappa-B, triggering synthesis of inflammatory mediators. Synergizes with IL12/interleukin-12 to induce IFNG synthesis from T-helper 1 (Th1) cells and natural killer (NK) cells.
-
基因功能參考文獻:
- This study identifies a role for IL18 in suppressing aberrant neuronal transmission in AD. PMID: 30127003
- Chimeric antigen receptor T cells releasing IL-18 convert to T-Bet(high) FoxO1(low) effectors that exhibit augmented activity against advanced solid tumors. PMID: 29241547
- The data suggest that inflammasome signaling is largely protective during murine coronavirus infection, in large part due to the pro-inflammatory effects of IL-18. PMID: 28895072
- downstream mediator of IL18 receptor activation, phospho-NF-kB, was increased in basolateral amygdala neurons expressing IL18 receptors PMID: 27590137
- This study provides the first evidence of the ability of IL-18 to induce B-type natriuretic peptide synthesis in vitro and outlines the relationship between the two molecules in acute HF patients with an ongoing inflammatory status. PMID: 24585936
- Interleukin-18 plays a role in advancing sepsis-induced cardiac dysfunction through inhibition of PP2A activity. PMID: 28526544
- These observations suggest that IL-18 exerts direct effects upon the GnRH neuron via IL-18Ralpha and acts on GnRH neurons through an autocrine or paracrine pathway. PMID: 28373090
- Reduced IL-18 serum concentration in children after HUS with no difference in its urine concentration may indicate a loss of the protective effects of this cytokine on renal function due to previously occurred HUS. PMID: 27982687
- In conclusion, liver X receptor activation inhibits IL-18 production through regulation of its transcription and maturation into an active pro-inflammatory cytokine. PMID: 27149934
- MyD88 signaling in myeloid and dendritic cells is dispensable for IFN-gamma-dependent control of type A F. tularensis infection. PMID: 28951422
- study found that IL-18 and IL-1beta are differentially regulated. Despite being constitutively expressed, IL-18 expression was increased and sustained after stimulation of TLRs. In contrast, IL-1beta was induced but not sustained after chronic treatment. PMID: 28468974
- This provides new insight into the immune phenotype, mechanisms, and signaling pathways that operate in microglial neurotoxic activation in amyotrophic lateral sclerosis. PMID: 28336525
- the GLA-SE adjuvant operates through interaction with IL-18-producing SCMsmall ef, Cyrillic for the rapid induction of B cell expansion and differentiation, Ab secretion, and Th1 responses PMID: 27794001
- IL-18 is essentially involved in mediating C. jejuni infection in the gnotobiotic mouse model. PMID: 27322540
- These results demonstrate a central role for the AIM2 inflammasome in preventing dysbiosis and intestinal inflammation through regulation of the IL-18/IL-22BP/IL-22 and STAT3 pathway PMID: 27524110
- Aldosterone induced IL-18 gene expression in renal tubular epithelial cells in a concentration- and time-dependent manner. PMID: 27413021
- this study demonstrated the critical function of IL-18 in lipid metabolism and these findings might contribute to the progress of novel treatments for nonalcoholic fatty liver disease or nonalcoholic steatohepatitis. PMID: 27063959
- Results indicated that IL-18 has roles apart from those as a proinflammatory cytokine in cardiac myocytes and suggested that IL-18 contributes to the homeostatic maintenance of mitochondrial function and gap-junction turnover in cardiac myocytes, possibly by upregulating autophagy. PMID: 27288439
- IL-18-elicited NK cell perforin responses seem to be critical for coordinating mucosal inflammation during early infection PMID: 27341123
- Data indicate that NLRC4 activation in Intestinal epithelial cells (IECs) leads to cell expulsion and IL-18 release, and implicate Caspase-8 in NLRC4 inflammasome responses in vivo by generation of doubly deficient in Caspase-1 and Caspase-8. PMID: 28410991
- this study shows that IL-18, cooperatively with IL-23, induced prominent inflammation and enhanced psoriasis-like epidermal hyperplasia PMID: 28299442
- telmisartan can reduce traumatic cerebral edema by inhibiting the NLRP3 inflammasome-regulated IL-1beta and IL-18 accumulation. PMID: 27465336
- Data, including data from studies in knockout mice, suggest that TNF (tumor necrosis factor), TNFR1 (TNF receptor 1), and IL18 (interleukin 18) are involved in liver lipid metabolism. IL18 or TNFR1 knockout mice on chow show higher liver triglyceride deposition than wild-type mice fed chow; in IL18 or TNFR1 knockout mice, high-refined-carbohydrate diet does not lead to fatty liver disease as it does in wild-type mice. PMID: 27863204
- GABA-B receptor antagonists reduced Il18 levels in male mice and increased Il18 levels in female mice following neonatal hypoxia ischemia. PMID: 27731803
- NLRP1 is an innate immune sensor that functions in the context of metabolic stress to produce IL-18, preventing obesity and metabolic syndrome. PMID: 26603191
- an important role in iNKT cell-mediated experimental eosinophilic esophagitis PMID: 25801352
- IL-18 is sufficient to protect Birc3-deficient mice from sodium dodecyl sulfate-induced colitis. PMID: 26037070
- IL-18 plays a facilitative role via NF-kappaB activation in pulmonary hypertension formation. PMID: 26440469
- IL-18 is a key epithelial-derived cytokine that differentially regulates distinct subsets of intestinal CD4(+) T cells during both homeostatic and inflammatory conditions PMID: 25736457
- Genetic ablation of IL-18 does not protect mice against maladaptive right ventricular remodeling following exposure to hypobaric hypoxia. PMID: 26747780
- We conclude that in the systemic lupus erythematosus syndrom IL-18 is involved specifically in the renal pathogenesis PMID: 26465326
- These results uncover the direct role of IL-18 in promoting goblet cell dysfunction during colitis, leading to breakdown of the mucosal barrier. PMID: 26638073
- IL-18 is a potent stimulator of Chi3l1 and that Chi3l1 is an important mediator of IL-18-induced inflammatory, fibrotic, alveolar remodeling, and cytotoxic responses. PMID: 25955511
- macrophages are not involved in IL-18-mediated susceptibility to L. amazonensis. PMID: 26009021
- The published data prompt to the hypothesis that IL-18 induces a broad spectrum of COPD-like inflammatory and remodeling responses in the murine lung PMID: 25922275
- Following hypoxic ischemic insult, long term creatine monohydrate supplementation up regulates the IL-6 and IL-18 concentrations triggering the neuroinflammatory and neuroprotective responses. PMID: 26639507
- High and fluctuating levels of glucose may be associated with inflammation and diabetic atherosclerosis by regulating the expression levels of IL-18. PMID: 25955000
- a new mechanism by which PKC-beta activation promotes EC dysfunction caused by the de-regulation of the IL-18/IL-18BP pathway, leading to increased VCAM-1 expression, monocyte/macrophage adhesion, and accelerated atherosclerotic plaque formation in diabetes PMID: 25808972
- PMA treatment during a vulnerable period can alter brain development. IL-18 and IRAK-4 appear to be important for the development of PMA induced injury. PMID: 25918710
- IL-18 levels modulate innate immune responses and cryptosporidiosis in mice. PMID: 25155632
- these results indicate that the TLR2/NLRP3/CASP1/IL-18 axis is critical to H. pylori-specific immune regulation. PMID: 26214524
- This study identifies NCC as an IL18-binding protein that collaborates with IL18r in cell signaling, inflammatory molecule expression, and experimental atherogenesis. PMID: 26099046
- Neutralization of IL-18 during alveolar hypoxia improves left ventricle diastolic function and partly prevents right ventricle hypertrophy PMID: 25182570
- This study reveals essential role of autophagy as a negative regulator of lung inflammation and identifies IL-18 as a critical mediator in lung injury due to autophagy deficiency. PMID: 25888640
- IL-18, but not IL-1beta has a role in aberrant actin depolymerization that triggers the pyrin inflammasome and autoinflammatory disease PMID: 26008898
- findings show how TLR2 deficiency accelerates IL18-mediated immunosuppression during liver carcinogenesis PMID: 25600646
- Nlrp3 inflammasome is essential for resistance against P. brasiliensis because it orchestrates robust caspase-1 activation and triggers an IL-18-dependent proinflammatory response. PMID: 25825440
- IL-18 can positively impact bone marrow lymphopoiesis and T cell development, presumably via interaction with the c-Kit and IL-7 signaling axis. PMID: 25780034
- Alprostadil treatment can protect renal function by reducing proteinuria. These effects are mediated, at least in part, through down-regulation of Ang-2 and IL-18 expression PMID: 25200363
- IL-18 was required for the expression of IL-22 in innate lymphoid cells upon Toxoplasma gondii infection. PMID: 25680273
顯示更多
收起更多
-
亞細胞定位:Cytoplasm. Secreted.
-
蛋白家族:IL-1 family
-
數據庫鏈接:
Most popular with customers
-
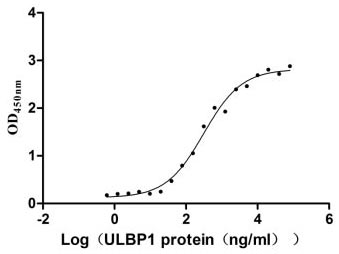
Recombinant Human UL16-binding protein 1 (ULBP1) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
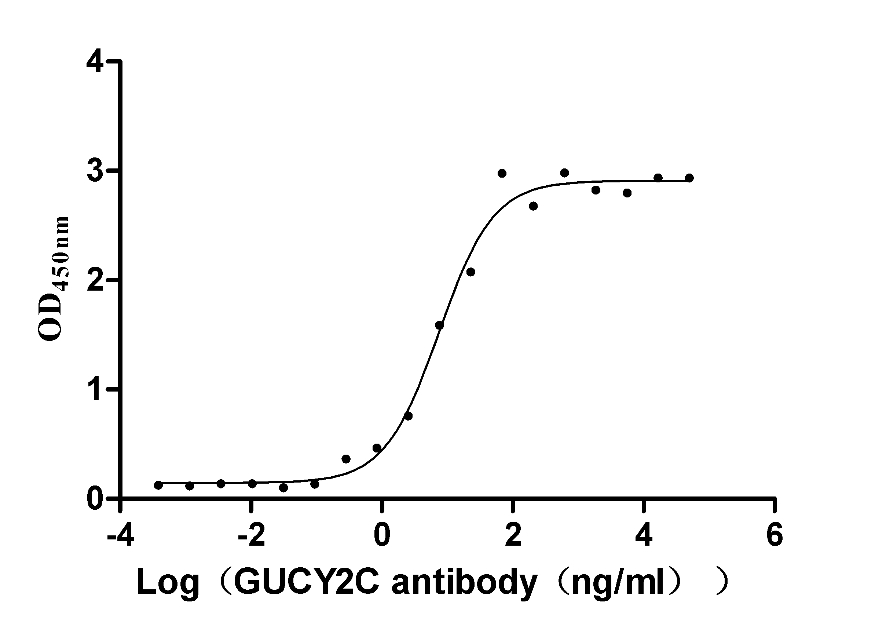
Recombinant Human Heat-stable enterotoxin receptor (GUCY2C), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
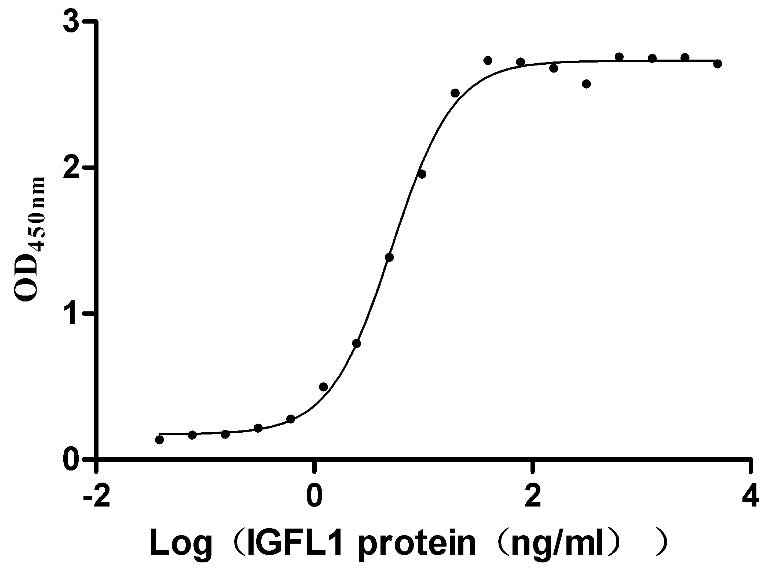
Recombinant Human IGF-like family receptor 1 (IGFLR1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
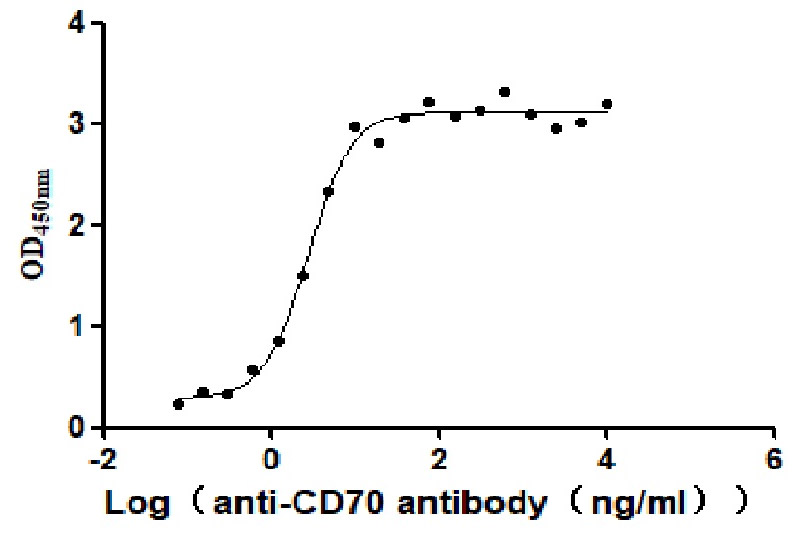
Recombinant Human CD70 antigen (CD70), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
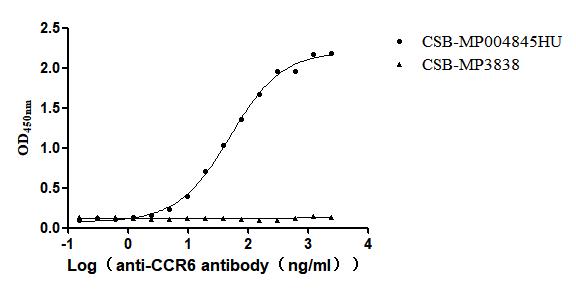
Recombinant Human C-C chemokine receptor type 6(CCR6)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
-
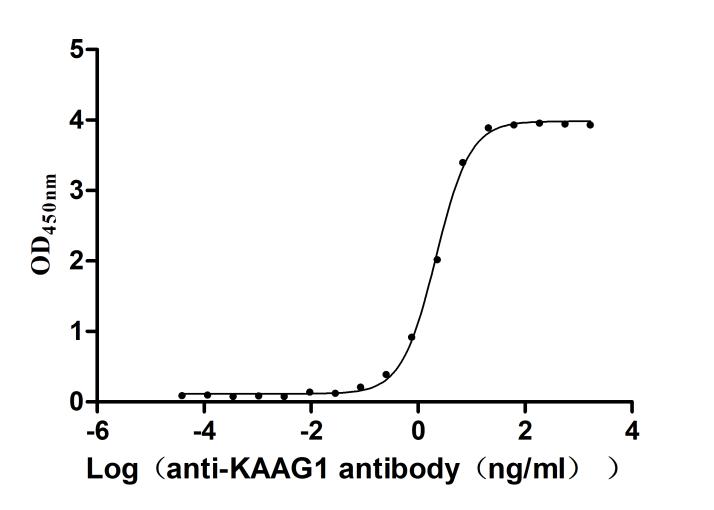
Recombinant Human Kidney-associated antigen 1 (KAAG1) (Active)
Express system: E.coli
Species: Homo sapiens (Human)
-
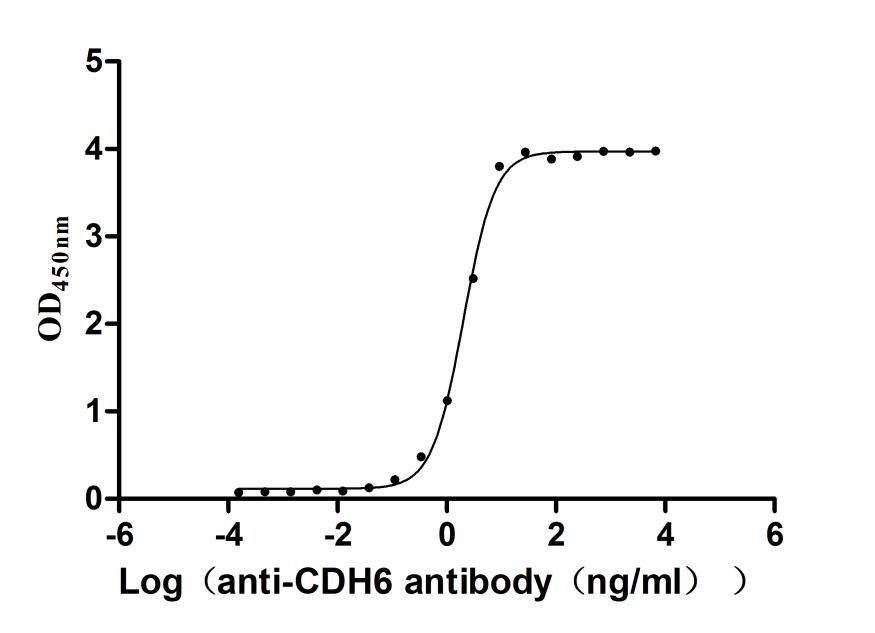
Recombinant Mouse Cadherin-6 (Cdh6), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)











