Recombinant Mouse Interleukin-13 (Il13), partial (Active)
In Stock-
中文名稱:小鼠Il13重組蛋白
-
品名簡稱:Recombinant Mouse Il13 protein, partial (Active)
-
貨號:CSB-AP004771MO
-
說明書:
-
規(guī)格:¥1740
-
圖片:
-
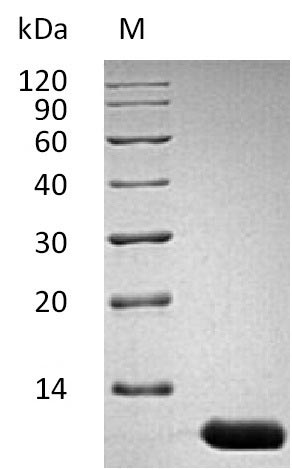 (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
-
-
其他:
產(chǎn)品詳情
-
純度:Greater than 95% as determined by SDS-PAGE.
-
內(nèi)毒素:Less than 1.0 EU/μg as determined by LAL method.
-
生物活性:The ED50 as determined in a cell proliferation assay using TF-1 human erythroleukemic cells is 1.93 ng/ml.
-
基因名:
-
Uniprot No.:
-
別名:Il13; Il-13; Interleukin-13; IL-13; T-cell activation protein P600
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Partial
-
來源:E.coli
-
分子量:11.7 kDa
-
表達(dá)區(qū)域:26-131aa
-
氨基酸序列SVSLPLTLKELIEELSNITQDQTPLCNGSMVWSVDLAAGGFCVALDSLTNISNCNAIYRTQRILHGLCNRKAPTTVSSLPDTKIEVAHFITKLLSYTKQLFRHGPF
-
蛋白標(biāo)簽:Tag-Free
-
產(chǎn)品提供形式:Liquid or Lyophilized powder
-
緩沖液:Lyophilized from a 0.2 μm filtered 20mM Histidine-HCl, 8% Trehalose, 4% Mannitol, 50mM NaCl, 0.05% Tween 80, pH 6.0.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Basically, we can dispatch the products out in 1-3 working days after receiving your orders. Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
Datasheet & COA:Please contact us to get it.
產(chǎn)品評價

相關(guān)產(chǎn)品
靶點詳情
-
功能:Cytokine. Inhibits inflammatory cytokine production. Synergizes with IL2 in regulating interferon-gamma synthesis. May be critical in regulating inflammatory and immune responses. Positively regulates IL31RA expression in macrophages.
-
基因功能參考文獻(xiàn):
- S1PR2 facilitates lung fibrosis through the mechanisms involving augmentation of IL-13 expression and its signaling in BALF cells. PMID: 29782549
- Combined blockade of the IL-13 and IL-33 pathways leads to a greater inhibition of type 2 inflammation over inhibition of either pathway alone. PMID: 27697499
- Both pre- and post-transcriptional processes may be involved in the AR modulation of ILC2 IL-5 and IL-13 production. PMID: 28982732
- the endothelial barrier was preserved in respiratory epithelium isolated from MCU-/- mice after exposure to IL-13. In the ovalbumin-model of allergic airway disease, MCU deficiency resulted in decreased apoptosis within the large airway epithelial cells. Concordantly, expression of the tight junction protein ZO-1 was preserved, indicative of maintenance of epithelial barrier function PMID: 29225050
- controls the rate of epithelial cell movement through the epidermis and acts as a molecular bridge between intraepithelial lymphocytes and epithelial cells PMID: 27357235
- results demonstrate that IL-13 is a major regulator of radiation-induced lung injury and demonstrates that strategies focusing on IL-13 may be useful in screening for timely delivery of anti-IL-13 therapeutics. PMID: 28004808
- Using a mouse model of Th2-mediated inflammation induced by OVA-allergen, this study observed elevated lung amounts of IL-13 and IL-4 accompanied by increased autophagosome levels, determined by LC3BII protein levels and immunostaining. PMID: 28982074
- Metaplasia induction and macrophage polarisation after parietal cell loss is coordinated through a cytokine signalling network of IL-33 and IL-13, linking a combined response to injury by both intrinsic mucosal mechanisms and infiltrating M2 macrophages. PMID: 28196875
- IL-13 is able to signal independent of the IL-4Ra chain in AD (atopic dermatitis), which may lead to the identification of molecular pathways downstream of IL-13 signaling that could be targeted in future therapies for AD. PMID: 26896776
- the presence of interleukin-13 (IL-13), which can convert inflammatory into Ym1+ alternatively activated macrophages, at (acinar-to-ductal metaplasia [ADM]), which then gives rise to pancreatic intraepithelial neoplasia lesions, is reported. PMID: 28514653
- Data indicate that interleukin-33 (IL-33)-induced Interleukin-13 (IL-13) production by type-2 helper T cells (Th2 cells) Is dependent on epidermal growth factor receptor (EGFR) expression. PMID: 29045902
- this study shows that environmental IL-13 plays a role in conditioning early thymic progenitors lineage choice, which would impact T cell development PMID: 28893952
- IL-4 and IL-13 are required to effectively polarize macrophages/dendritic cells to an M2a phenotype and to promote recovery from acute kidney injury. PMID: 27745702
- this study shows that ST2 regulates early IL-13 production in fungus-induced allergic airway inflammation PMID: 26555705
- These observations suggest that IL-4 and IL-13 likely operate through the Heteroreceptor and influence Th17 cells to convert to Th1 cells and to acquire increased sensitivity to suppression, leading to control of immune-mediated CNS inflammation. PMID: 28801358
- MIF-deficient mice have reduced Nippostrongylus brasiliensis burden and mounted an enhanced type 2 immune response, including increased Gata3 expression and IL-13 production in the mesenteric lymph nodes PMID: 27049059
- findings suggest that a leukotriene B4 receptor-2-linked cascade plays a pivotal role in LPS/TLR4 signaling for IL-13 synthesis in mast cells, thereby potentially exacerbating allergic response. PMID: 28600286
- Study found IL-13 to be critically involved in the development of chemical-induced asthma, as shown by using IL-13 KO mice, and more specifically in the effector phase as confirmed by anti- IL-13 antibody treatment. PMID: 28704401
- these studies show that fibrosis, steatosis, cholestasis, and ductular reaction are simultaneously controlled but distinctly regulated by interleukin-13 signaling PMID: 27421703
- Our data support that impaired clearance of inhaled allergens triggering IL-13 production by multiple cell types in the airways plays an important role in the pathogenesis of type 2 airway inflammation and suggests therapeutic improvement of mucociliary clearance as a novel treatment strategy for children with allergen-induced asthma. PMID: 27865862
- this study shows that wild-type mice develop an eosinophilic Th2 airway disease in response to Alternaria alternata exposure, whereas IL-13-deficient mice exhibit a primarily neutrophilic response PMID: 27815425
- this study shows that IL-17A contributes to asthma pathophysiology by increasing the capacity of IL-13 to activate intracellular signaling pathways, such as STAT6 activation PMID: 27417023
- RCM-1 reduced IL-13 and STAT6 (signal transducer and activator of transcription 6) signaling and prevented the expression of the STAT6 target genes Spdef and Foxa3, which are key transcriptional regulators of goblet cell differentiation. PMID: 28420758
- IL-13 suppressed both the activation-induced apoptosis of CD4(+) T cells and the expression of p53 and FasL. PMID: 26189367
- We clearly show that miR-155 has a previously unknown direct regulatory role in the ILC2 subset that affects IL-33 receptor expression, IL-33 responsiveness, and IL-13 production as well as proliferation capability, possibly due to defects in GATA-3 function. PMID: 27492144
- The presented data substantiate the hypothesis that claudin-18 is a central barrier-forming component of tight junctions and show that IL-13 downregulates claudin-18. These data also suggest that the loss of claudin-18 is associated with increased sensitization to aeroantigens and airway responsiveness PMID: 27215490
- Studies in colonic T84 cell monolayers revealed that barrier disruption by the colitis-associated Th2-type cytokines, IL-4 and IL-13, down-regulates matriptase as well as prostasin through phosphorylation of the transcriptional regulator STAT6 PMID: 28490634
- These data demonstrate that multiple pathogenic strains of RSV induce IL-13-producing group 2 innate lymphoid cell proliferation and activation through a TSLP-dependent mechanism in a murine model and suggest the potential therapeutic targeting of TSLP during severe RSV infection. PMID: 27156176
- The soluble antigen from A. cantonensis could promote the Chil3 expression in macrophage and microglial cell lines induced by interleukin-13. PMID: 27256220
- The reduction in fibrosis observed when IL-13 signalling is suppressed is not dependent on increased IFN-gamma activity. Instead, by reducing compensatory increases in type 1-associated inflammation, therapeutic strategies that block IFN-gamma and IL-13 activity simultaneously can confer greater protection from progressive fibrosis than IL-13 blockade alone. PMID: 27125685
- The IL-23/IL-17 axis plays a critical role in the immunopathology of hepatic amebiasis. IL-13 secreted by CD11b(+)Ly6C(lo) monocytes may be associated with recovery from liver damage. PMID: 26809113
- PLD1 activation enhanced binding of ROCK1 to ATF-2 and leads to increased expression of IL-13 PMID: 26335962
- Macrophages are critical to the maintenance of IL-13-dependent lung inflammation and fibrosis. PMID: 25921340
- IL-25 and CD4(+) TH2 cells enhance type 2 innate lymphoid cell-derived IL-13 production, which promotes IgE-mediated experimental food allergy. PMID: 26560039
- Placenta growth factor augments airway hyperresponsiveness via leukotrienes and IL-13. PMID: 26690703
- Natural helper cells contribute to pulmonary eosinophilia by producing IL-13 via IL-33/ST2 pathway in a murine model of respiratory syncytial virus infection PMID: 26044350
- review of IL-4 and IL-13 mast cell immunity and detail of the differences that exist between mouse and human mast cell responses to IL-4 and IL-13 [review] PMID: 26088754
- Data (including data from studies in knockout/transgenic mice) suggest T cell-derived IL4/IL13 are required for immunologic memory and IgE response to helminth Nippostrongylus brasiliensis but are not required for expansion/proliferation of B cells. PMID: 26523376
- Curcumin up-regulates mRNA and protein levels of IL-4 and IL-13 PMID: 25944087
- These data indicate that distal airways might be less sensitive to IL-13-induced GC metaplasia and mucus production through lower expression of IL-13Ralpha1 and attenuated activation of downstream signalling. PMID: 25772331
- IL-13 induces miR-142-5p and downregulates miR-130a-3p in macrophages, regulating macrophage profibrogenic gene expression in chronic inflammation. PMID: 26436920
- IL-4 and IL-13 have a critical role in innate immune cells for protective immunity against gastrointestinal helminths. PMID: 25336167
- These data demonstrate that dysregulated IL-25 expression contributes to lipid accumulation, whereas exogenous IL-25 protects against hepatic steatosis through IL-13 activation of STAT6. PMID: 26423151
- TH2 cells and their cytokines IL-4 and IL-13 regulate formation and function of lymphatic vessels. PMID: 25648335
- Mice with experimental Schistosoma-induced pulmonary hypertension (PH) had evidence of increased IL-4 and IL-13 signaling. IL-4(-/-)IL-13(-/-) mice, but not single knockout IL-4(-/-) or IL-13(-/-) mice, were protected from Schistosoma-induced PH. PMID: 26192556
- regulates the expression of IL-17A in HIV-specific CD8 T cells following immunizations PMID: 25493691
- These data establish for the first time a molecular mechanism by which Mac-1 regulates the signaling activity of IL-13 in macrophages. PMID: 26160172
- Acidic pH augments Fc-epsilon-RI-mediated production of IL-6 and IL-13 in mast cells. PMID: 26196745
- conjunctival goblet cells are IL-13 responsive cells that produce factors known to maintain epithelial barrier, stimulate mucin production, and modulate immune response in nonocular mucosa when treated with IL-13. PMID: 26132778
- Enhanced IL-13 production by T cells can play a causative role in the exocrinopathy observed in Id3 knockout mice. PMID: 25010390
顯示更多
收起更多
-
亞細(xì)胞定位:Secreted.
-
蛋白家族:IL-4/IL-13 family
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-
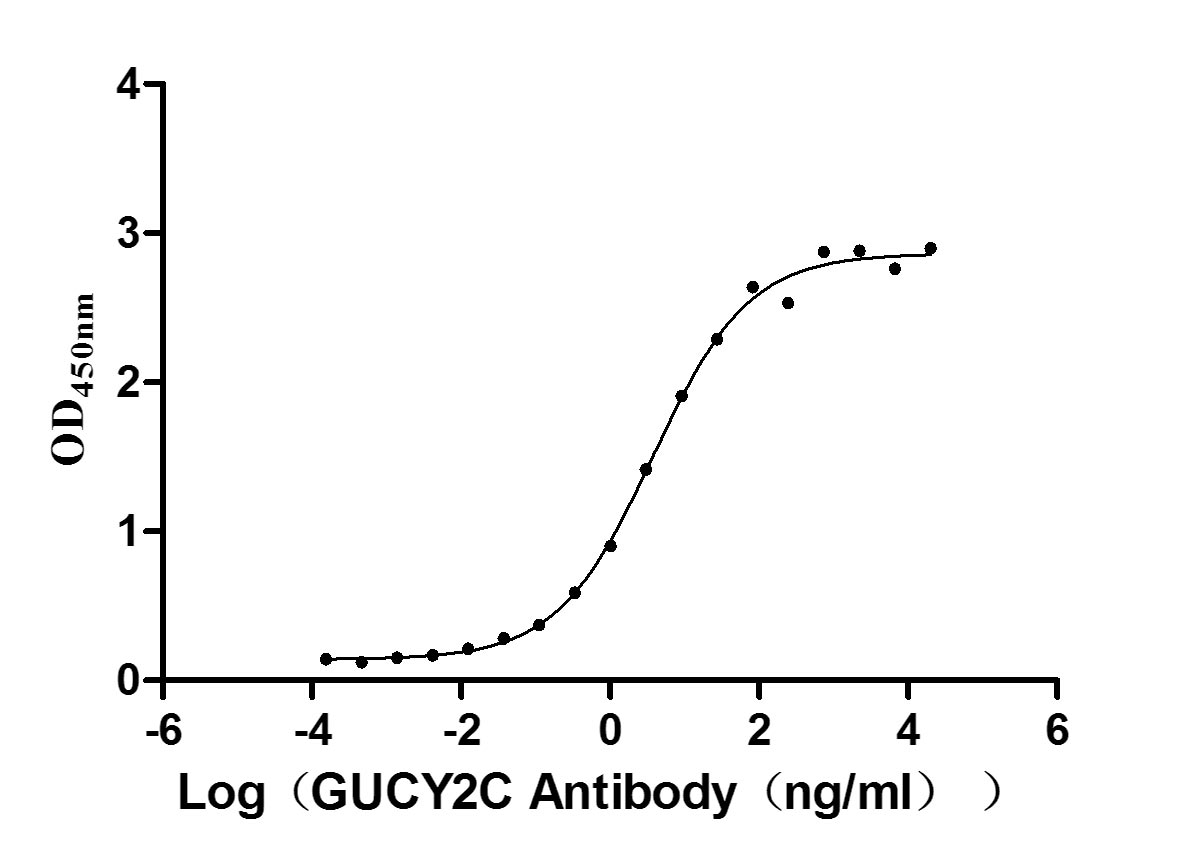
Recombinant Human Heat-stable enterotoxin receptor (GUCY2C), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
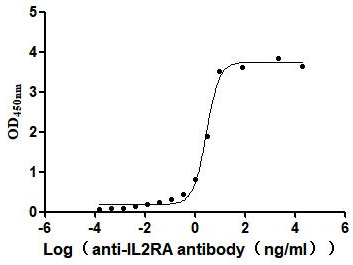
Recombinant Human Interleukin-2 receptor subunit alpha (IL2RA), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
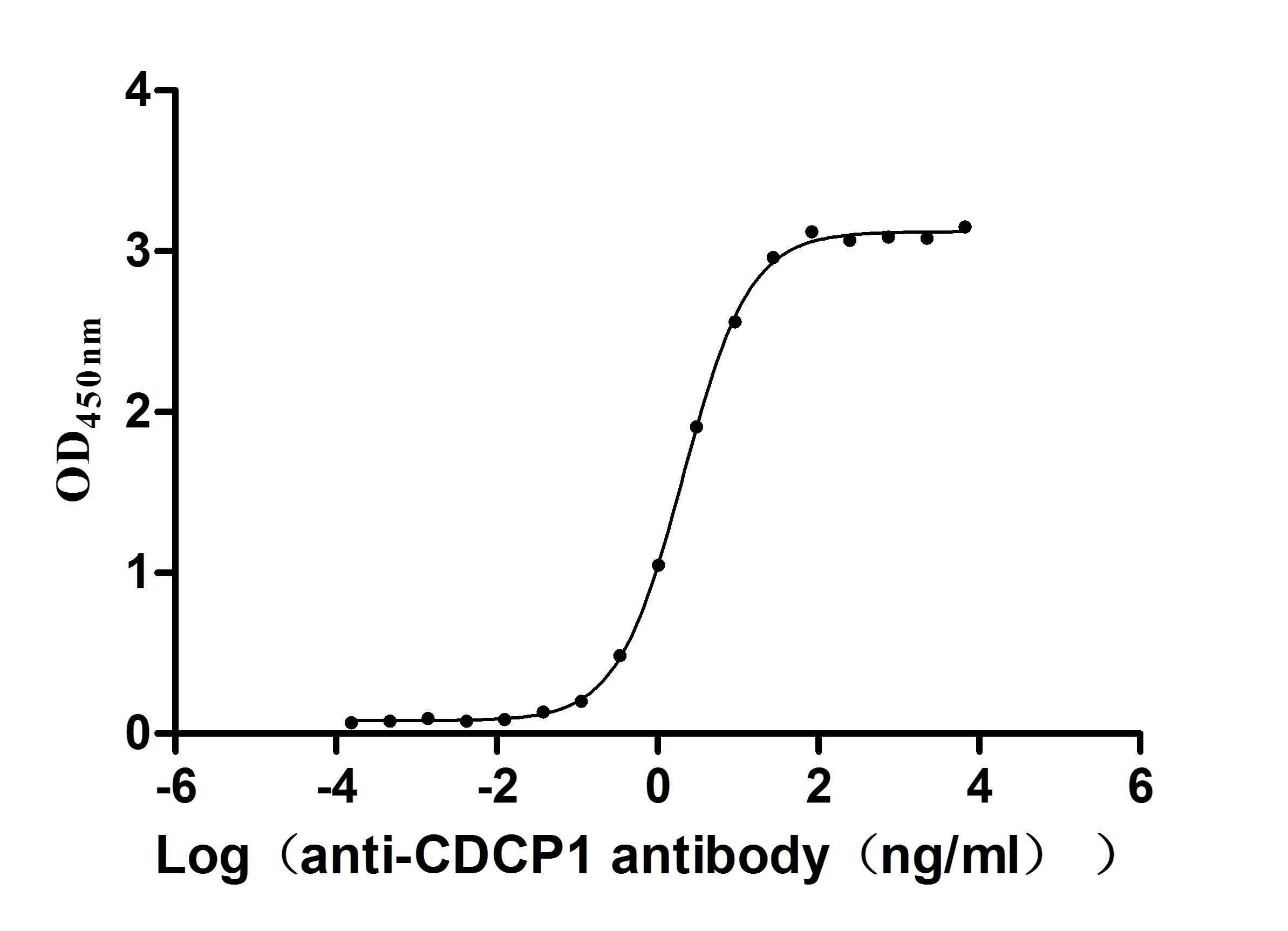
Recombinant Macaca fascicularis CUB domain containing protein 1 (CDCP1), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant Human Cadherin-1(CDH1),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)











