Recombinant Mouse Granulocyte-macrophage colony-stimulating factor protein (Csf2)
In Stock-
中文名稱:Recombinant Mouse Granulocyte-macrophage colony-stimulating factor protein(Csf2) (Active)
-
品名簡稱:Recombinant Mouse Csf2 protein (Active)
-
貨號:CSB-AP003441MO
-
說明書:
-
規格:¥852
-
圖片:
-
-
其他:
產品詳情
-
純度:>98% as determined by SDS-PAGE.
-
內毒素:Less than 1.0 EU/μg as determined by LAL method.
-
生物活性:Fully biologically active when compared to standard. The ED50 as determined by a cell proliferation assay using murine FDC-P1 cells is less than 0.05 ng/ml, corresponding to a specific activity of >2.0x107 IU/mg.
-
基因名:
-
Uniprot No.:
-
別名:Csf2; Csfgm; Granulocyte-macrophage colony-stimulating factor; GM-CSF; Colony-stimulating factor; CSF
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Full Length of Mature Protein
-
來源:E.Coli
-
分子量:14.1 kDa
-
表達區域:18-141aa
-
氨基酸序列APTRSPITVT RPWKHVEAIK EALNLLDDMP VTLNEEVEVV SNEFSFKKLT CVQTRLKIFE QGLRGNFTKL KGALNMTASY YQTYCPPTPE TDCETQVTTY ADFIDSLKTF LTDIPFECKK PGQK
-
蛋白標簽:Tag-Free
-
產品提供形式:Liquid or Lyophilized powder
-
緩沖液:0.2μm filtered PBS, pH 7.4 ,lyophilized
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:5-10 business days
-
Datasheet & COA:Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:Cytokine that stimulates the growth and differentiation of hematopoietic precursor cells from various lineages, including granulocytes, macrophages, eosinophils and erythrocytes.
-
基因功能參考文獻:
- Data show that the microbiota enhances respiratory defenses via granulocyte-macrophage colony-stimulating factor (GM-CSF) signaling, which stimulates pathogen killing and clearance by alveolar macrophages PMID: 29142211
- The data indicate that GM-CSF drives chronic tissue damage and disability in experimental autoimmune encephalomyelitis via pleiotropic pathways, but it is dispensable during early lesion formation and the onset of neurologic deficits. PMID: 29288202
- the release of IL-33 and GM-CSF from epithelial cells induces the activation of p65 and the p38-MK2/3 signaling module in Dendritic Cells, resulting in Th2 polarization and, finally, allergic inflammation. PMID: 29288203
- results show T cell production of GM-CSF contributes to control of M. tuberculosis infection in the absence of other sources of GM-CSF, that multiple T cell subsets make GM-CSF in the lung over the course of infection and that GM-CSF can act directly on infected macrophages through a pathway requiring PPARgamma to limit bacterial growth PMID: 29066547
- In conclusion, our study confirms the pathogenic role of GM-CSF in colitis-associated colorectal cancer development. GM-CSF favors tumor-permissive microenvironment by inducing MDSC generation and recruiting them into colonic tissues. PMID: 28534709
- these data demonstrate that GM-CSF levels during radiotherapy can be used as a prognostic biomarker for lung and esophageal cancer PMID: 27835886
- this study demonstrates that epithelial-derived GM-CSF is a critical early signal during allergic sensitization to cockroach allergen PMID: 27731325
- These impaired macrophage functions in leukemic mice were significantly corrected by IL-3 and GM-CSF treatment indicating the therapeutic benefit of these two cytokines in leukemia. PMID: 28039843
- Both IL-6 protein production and transcript levels were downregulated by RA in respiratory tract epithelial cells (LETs) , but upregulated in macrophages (MACs). RA also increased transcript levels of MCP-1, GMCSF, and IL-10 in MACs, but not in LETs. Conversely, when LETs, but not MACs, were exposed to RA PMID: 27940088
- T-GM-CSF and -IL-3 significantly, and reciprocally, blunted receptor binding and myeloid progenitor cell proliferation activity of both FL-GM-CSF and -IL-3 in vitro and in vivo PMID: 28344320
- Results indicate GM-CSF as both a key contributor to the pathogenesis of MI and a potential therapeutic target. PMID: 28978634
- Obesity alters the lung neutrophil infiltration to enhance breast cancer metastasis through IL5 and GM-CSF. PMID: 28737771
- GM-CSF is required for the normal balance of leukocyte subsets, including granulocytes, B cells, and naive vs. effector T cells. There was an approximately 3-fold increase in the percentages of granulocytes in Csf2-/- PBMCs. The presence of maximal experimental autoimmune encephalomyelitis in the complete absence of GM-CSF revealed that GM-CSF is not an obligate effector molecule in all forms of EAE. PMID: 27256565
- chemerin inhibited nuclear factor-kappaB activation and the expression of granulocyte-macrophage colony-stimulating factor (GM-CSF) and interleukin-2 (IL-6) by tumor cells and tumor-associated endothelial cell, respectively, via its receptors, and consequently, MDSC induction was impaired, leading to restoration of antitumor T-cell response and decreased tumor angiogenesis. PMID: 28166197
- These findings describe a novel role for GM-CSF as an essential initiating cytokine in cardiac inflammation. PMID: 27595596
- Data reviewed establish that any damage to brain tissue tends to cause an increase in G-CSF and/or GM-CSF (G(M)-CSF) synthesized by the brain. Glioblastoma cells themselves also synthesize G(M)-CSF. G(M)-CSF synthesized by brain due to damage by a growing tumor and by the tumor itself stimulates bone marrow to shift hematopoiesis toward granulocytic lineages away from lymphocytic lineages. PMID: 28459367
- Evi1(+)DA-3 cells modified to express an intracellular form of GM-CSF, acquired growth factor independence and transplantability and caused an overt leukemia in syngeneic hosts, without increasing serum GM-CSF levels. PMID: 25907616
- IL-23-induced GM-CSF mediates the pathogenicity of CD4(+) T cells in experimental autoimmune myocarditis. PMID: 26660726
- GM-CSF accelerated the G1/S phase transition in EPCs by upregulating the expression of cyclins D1 and E. PMID: 24662605
- host RNF13 affects the concentration of GM-CSF in tumor-bearing lungs PMID: 26197965
- Sc CW-derived BG stimulated the late and strong expression of Csf2 in a dectin-1-dependent manner, they remain poor inducers of chemokine and cytokine production in murine macrophages. PMID: 26840954
- GM-CSF and uPA are required for Porphyromonas gingivalis-induced alveolar bone loss in a mouse periodontitis model. PMID: 25753270
- These findings identify GM-CSF as central to the protective immune response that prevents progressive fungal disease PMID: 26755822
- regulatory molecule causative of aortic dissection and intramural haematoma PMID: 25923510
- Results indicate that granulocyte-macrophage colony-stimulating factor (GM-CSF) signaling controls a pathogenic expression signature in CC chemokine receptors 2 (CCR2+)Ly6C(hi) monocytes. PMID: 26341401
- TL1A increases expression of CD25, LFA-1, CD134 and CD154, and induces IL-22 and GM-CSF production from effector CD4 T-cells PMID: 25148371
- Mafb-deficient microglia cultured with GM-CSF barely extended their membrane protrusions, probably due to abnormal activation of RhoA, a key regulator of cytoskeletal remodeling. MafB is a negative regulator of GM-CSF signaling in microglia. PMID: 25998393
- Csf2-/- mice showed a 30% increase in whole body adiposity, which persisted to adulthood. Adipocytes from Csf2-/- mice were 50% larger by volume and contained higher levels of adipogenesis gene transcripts, indicating enhanced adipocyte differentiation. PMID: 25931125
- Renal ischemia reperfusion injury tubular cells expressed elevated GM-0CSF, that supports tubular cells proliferation. PMID: 25388222
- GM-CSF is key to the development of experimental osteoarthritis and its associated pain. PMID: 22995428
- Loss of encephalitogenic activity of STAT5-deficient autoreactive CD4+ T cells was independent of IFN-gamma or IL-17 production, but was due to the impaired expression of GM-CSF, a crucial mediator of T-cell pathogenicity. PMID: 25412660
- These data indicate that GM-CSF plays a role in the inflammatory signaling network that drives neutrophil recruitment in response to Clostridium difficile infection but does not appear to play a role in clearance of the infection. PMID: 25045999
- IRF8 represses GM-CSF expression in T cells to affect myeloid cell lineage differentiation. PMID: 25646302
- Reprogramming of monocytes by GM-CSF contributes to regulatory immune functions during intestinal inflammation. PMID: 25653427
- GM-CSF promotes advanced plaque progression by increasing macrophage apoptosis susceptibility. PMID: 25348165
- although iTreg cells maintained the ability to produce IFN-gamma and TNF-alpha in vivo, their ability to produce GM-CSF was selectively degraded upon antigen stimulation under inflammatory conditions. PMID: 25168419
- IFN-gamma has a pprotective role in the demyelination of brain through downregulation of IL-17/GM-CSF and induction of neuroprotective factors in the brain by monocytes/microglial cells. PMID: 25339670
- Expression of PPAR-gamma in fetal lung monocytes was dependent on the cytokine GM-CSF. Therefore, GM-CSF has a lung-specific role in the perinatal development of alveolar macrophages through the induction of PPAR-gamma in fetal monocytes. PMID: 25263125
- These novel findings indicate that the inflammatory mediator, GM-CSF, may have significant protective properties in the chronic sequelae of experimental Traumatic brain injury PMID: 24392832
- Airway infections and pollutants increase the susceptibility to develop allergic asthma through a GM-CSF/IL-33/OX40L pathway. PMID: 24551140
- Data indicate that neutralization of GM-CSF would be a useful therapeutic strategy for severe connective tissue diseases (CTD)-interstitial lung disease (ILD). PMID: 24951817
- Data indicate that lung epithelium-derived GM-CSF is a critical regulator of CD11b+ dendritic cells (DCs)-mediated Th2 cell priming. PMID: 24943219
- Bhlhe40 is required positively regulates the production of GM-CSF and negatively regulates the production of IL-10 in T cells. PMID: 24699451
- Report IL-18 plus GM-CSF modified tumor cell vaccine can induce significant antitumor effects in Lewis lung carcinoma. PMID: 24475975
- Data indicate that in type 1 diabetes monocytes STAT5Ptyr activation is significantly higher and is found bound to CSF2 promoter and PTGS2 enhancer regions. PMID: 24204704
- Data indicate that GM-CSF controls IgM production in innate response activator B cells. PMID: 24821911
- our findings suggest that over-production of GM-CSF by T cells could be a pathogenic factor in many histiocytic disorders PMID: 24183847
- that p52 binds to the promoter of the GM-CSF-encoding gene (Csf2) and cooperates with c-Rel in the transactivation of this target gene. PMID: 24899500
- GM-SCF, IL-21 and Rae1 expression, alone or in combination, induces a cellular immune response against H22 tumor cells. PMID: 24350772
- Data indicate that GM-CSF knockout mice exhibit a unique mix of M1-M2 macrophage phenotypes inalveolar macrophages. PMID: 24044676
顯示更多
收起更多
-
亞細胞定位:Secreted.
-
蛋白家族:GM-CSF family
-
數據庫鏈接:
Most popular with customers
-
Recombinant Human Tumor necrosis factor ligand superfamily member 14 (TNFSF14), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Heat-stable enterotoxin receptor (GUCY2C), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Tumor necrosis factor ligand superfamily member 9 (TNFSF9), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
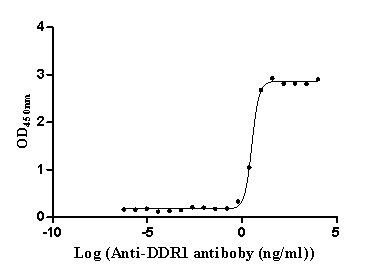
Recombinant Human Epithelial discoidin domain-containing receptor 1 (DDR1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
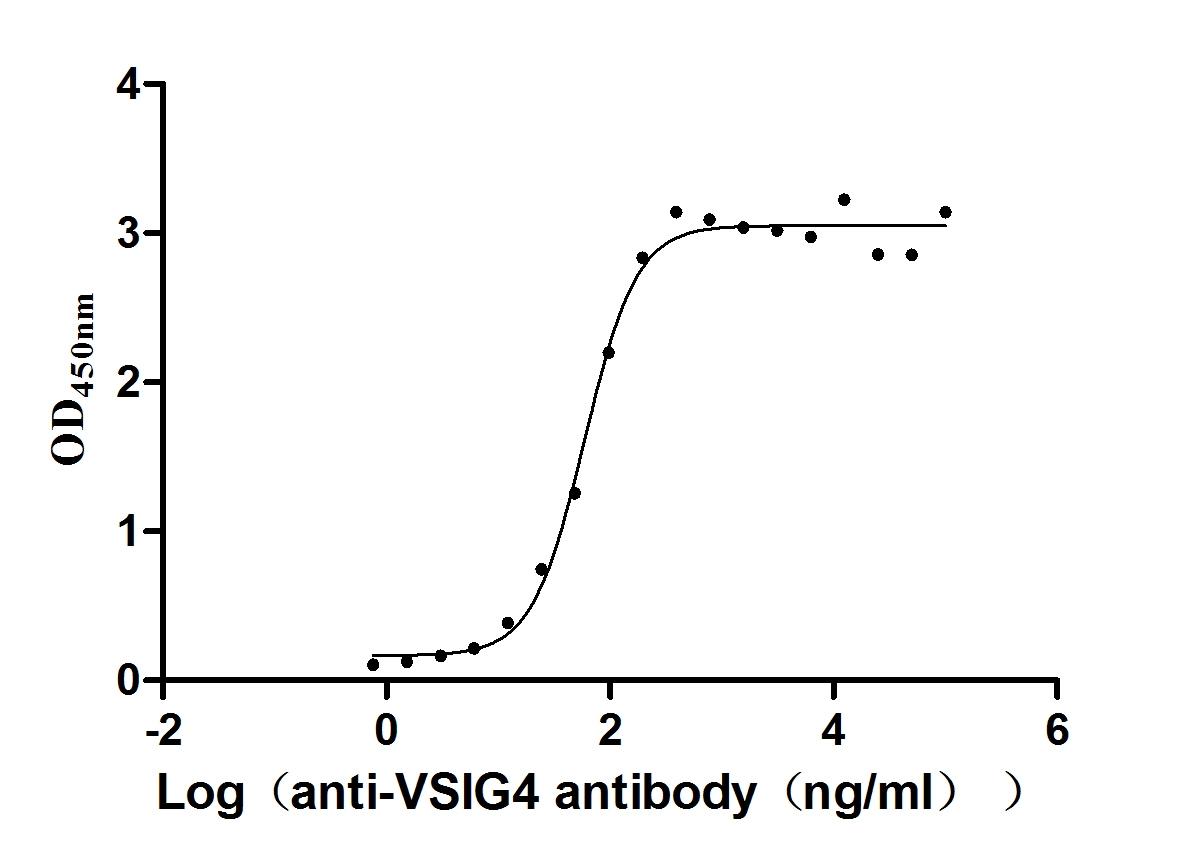
Recombinant Human V-set and immunoglobulin domain-containing protein 4 (VSIG4), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
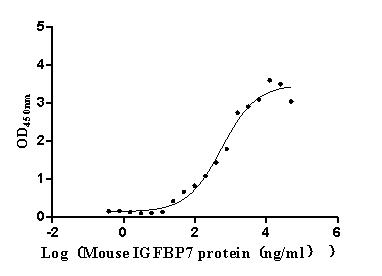
Recombinant Mouse Complement component C1q receptor (Cd93), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
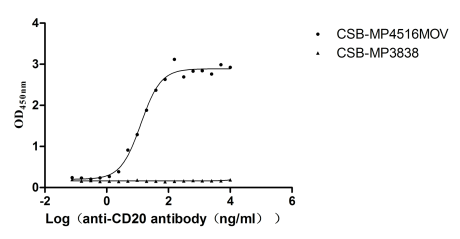
Recombinant Macaca fascicularis Membrane spanning 4-domains A1 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant Human Interleukin-2 receptor subunit alpha (IL2RA), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









